INTRODUCCIÓN
Inmunología es la rama de la medicina que estudia al sistema inmune. Este es el sistema encargado de la respuesta protectora frente a diversos microorganismos, la cual está dotada de especificidad y memoria. También el sistema inmune participa en el rechazo a trasplantes y en la protección frente a tumores.1
Hoy en día es conocido que, ante la exposición a un antígeno, el sistema inmune pude responder fundamentalmente de dos maneras: iniciar una respuesta inmune efectora la cual intentará eliminar la fuente antigénica cuando se trata de una antígeno foráneo, o responder con tolerancia cuando se trata de un antígeno propio.2 Ambas variantes son respuestas inmunes activas dotadas de especificidad y memoria cuyo fin es garantizar la permanencia del antígeno en nuestro organismo.
Los mecanismos de tolerancia evitan el desarrollo de procesos autoinmunes, y de respuestas innecesarias contra microorganismos comensales o no patogénicos.3 Los antígenos que inducen este estado de tolerancia se denominan tolerógenos. A diferencia de los que originan respuestas inmunes a los cuales se les denominan inmunógenos.4
El problema fundamental de la inmunología versa en torno a la anterior aseveración. Pues aun hoy en día no son del todo conocidos los mecanismos que intervienen en la “decisión” del sistema inmune de cuando responder con una respuesta fundamentalmente efectora y cuando hacerlo con una respuesta fundamentalmente tolerante.
A lo largo de la historia el pensamiento ha ido variando a medida que las tecnologías disponibles han permitido entender el funcionamiento del sistema inmune. Por esta razón se realizó la presente investigación bibliográfica con el propósito de resumir los cambios en el pensamiento inmunológico. Se realizó la revisión de artículos, muchos de ellos publicados en los últimos años. Para ello se hizo un búsqueda utilizando las expresiones “theory of inmune system” , “hstory of inmunology” y “inmunological tolerance”.
DESARROLLO
I.-Teorías clásicas del sistema inmune
- Horror autotoxicus
En 1901 Paul Erlich especuló sobre la aparición en algunos individuos de autoanticuerpos “tóxicos” y que estos estaban involucrados en la patogenia de algunos tipos de enfermedades.5 Erlich hace este planteamiento en el contexto de la teoría de las cadenas laterales, donde se especulaba que las células tenían cadenas laterales las cuales se podían unir a toxinas por un modelo de llave-cerradura. En condiciones de infección estas cadenas laterales se rompían y pasaban a ser los anticuerpos.6
El basamento de la teoría del horror antitoxicus se sustentaba en los experimentos de Bordet. Este científico demostró la aparición de anticuerpos antieritrocitarios que causaban hemólisis al inyectar glóbulos de un animal en otro y la imposibilidad de encontrar estos anticuerpos antieritrocitarios cuando los animales eran inmunizados con su propia sangre. De estas observaciones Erlich concluye que no se podían generar anticuerpos contra lo propio, dado a que se carecía de los receptores apropiados en las células para reconocerlos y, probablemente, cuando estos aparecían, eran suprimidos por su acción tóxica en el organismo.5
En 1955 Jerner plantea el punto de vista de Erlich pero con otras palabras, al decir que existe un repertorio preexistente de receptores del cual el antígeno hace una selección.7
A pesar de que hoy en día la teoría de las cadenas laterales ha sido completamente desechada, Erlich fue el primero en plantear la existencia del componente humoral de la inmunidad. También con su concepto de horros atutoxicus define una de las características del sistema inmune, como es la no reactividad frente a lo propio. Aunque este concepto hoy en día se trata desde un punto de vista diferente.
-La teoría de la selección clonal
La teoría más popular fue la de la selección clonal (TSC). Esta teoría fue desarrollada por Frank Macfarlane Burneo y colaboradores durante las décadas de 1940 y 1950, y ha sido fundamental para el avance de la inmunología contemporánea. La TSC, tal cual fue presentada por Burneo (1959), está formada por cuatro proposiciones principales:8 (Figura 1)
- El organismo tiene la capacidad innata para producir el universo de anticuerpos necesarios para responder a cualquier reto inmune.
- Cada anticuerpo es específico para un solo antígeno.
- Tras la estimulación por parte de su antígeno específico, las células portadoras de anticuerpo proliferarán para producir un conjunto de células exactamente iguales en especificidad a la célula madre.
- La antigenicidad es el resultado de un proceso de aprendizaje durante la vida embrionaria. Por tanto los linfocitos autorreactivos están ausentes del repertorio por ser eliminados en los estadios tempranos de la ontogenia.
La tesis central de la teoría de la selección clonal es la capacidad del sistema inmune para reconocer cualquier antígeno que pudiera estar presente en el organismo debido a una serie de mecanismos genéticos, somáticos, bioquímicos y celulares. Esto es consecuencia de la gran cantidad de células precursoras producidas, que tras la estimulación adecuada, originarán un conjunto de células efectoras específicas para el antígeno que provocó su producción (de ahí la selección clonal). Evidentemente tal capacidad debe tener ciertos límites a fin de evitar la autoinmunidad, para lo cual la teoría de la selección clonal propuso una serie de señales que regularían el sistema. Burnet mismo llamó a tales señales como propio y no propio.9
Esta teoría también incluye que el sistema inmune eliminará todas las células precursoras “antipropio”, donde propio es definido como el conjunto de todas las proteínas presentes en el individuo en algún momento de la vida embrionaria.10 Este postulado está basado en observaciones realizadas en la década de los 40.11
No es hasta la década de los 80 que se logra demostrar el fenómeno de delección clonal mediante el que son eliminados los clones autorreactivos como mecanismos de tolerancia central, se corrobora así la genialidad de Burnet al plantear esta teoría tres décadas antes.12 Como limitantes de esta teoría es que nos dice que los clones solo responden ante antígenos no propios, por lo que existe un estado de no respuesta frente a lo propio (tolerancia como fenómeno pasivo), existe además ausencia de clones autorreactivos en periferia y los receptores de las células efectoras son idénticos a los del clon que les dio origen; por lo que niega los fenómenos de maduración de la afinidad donde también pueden aparecer clones autorreactivos.
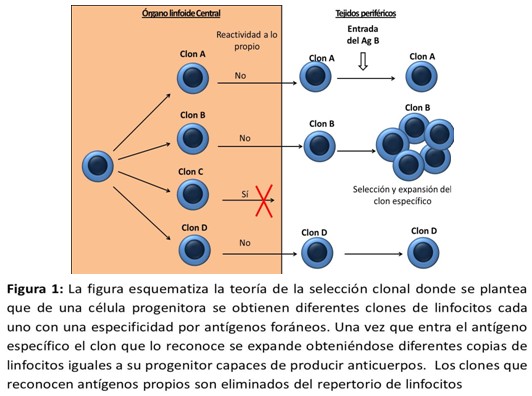
Dentro de los principales aciertos de la teoría de la selección clonal está el describir cómo es que ocurre una respuesta inmune antígeno específica. También explica en cierta medida como operan los mecanismos de tolerancia central, sin embargo hoy en día se sabe que no todos los linfocitos que reconocen lo propio son eliminados, pues una parte pasan a periferia y están sujetos a mecanismos de tolerancia periférica. A pesar de esto hay que reconocer la genialidad de Burnet el cual enuncia su teoría aun sin existir tecnología que le permitieran probarlo.
-La teoría de lo propio y lo no propio
El concepto de propio y no propio fue ampliamente utilizado en la década de los 60; el propio Brunet en conferencias dadas en la universidad de Cambridge abordó estos conceptos. En general estos conceptos refuerzan la idea de la selección clonal donde se habla del “estado de no respuesta contra lo propio”,13 siendo así la tolerancia un fenómeno pasivo inherente al sistema inmune y cualquier manifestación de autoinmunidad sería considerado como un fallo de este mecanismo fisiológico.14,15
Inicialmente se definió como propio al conjunto de proteínas expresadas durante la vida embrionaria. Sin embargo la falta de respuesta frente a la microbiota y la expresión de antígenos posteriores al nacimiento frente a los cuales “no existía” respuesta inmune llevo a rediseñar el concepto de propio como aquel conjunto de antígenos altamente expresado en el individuo desde edades tempranas de la vida.16
Lo significativo de estos conceptos es que trataban de explicar las cualidades que debería reunir un antígeno para ser tolerado, en otras palabras, se estaba definiendo qué era lo propio. Sin embargo el gran aporte de estos conceptos es que por primera vez se acepta la existencia de un reconocimiento por parte del sistema inmune de lo propio. O sea que el sistema inmune censaba lo propio para, de alguna manera, no responder ante ello.
-La teoría de las dos señales
En la práctica experimental fue detectada autorreactividad frente a lo propio, sin embargo no se evidenció enfermedad, por tanto debían existir mecanismos en periferia que regularan la capacidad de estos clones de activarse frente a antígenos propios; por tanto de alguna manera debía existir una tolerancia en periferia que controlara estos clones autorreactivos.17
La teoría de la selección clonal no podía explicar estos hallazgos, por lo que en 1972 Bretcher y Chon postulan la teoría de las dos señales, la cual en esencia plantea que para lograr la completa activación de las células se necesitaban dos señales, la primera proveniente del reconocimiento del antígeno a través de los receptores y una segunda señal que dependía de interacciones con la célula presentadora de antígenos. En ausencia de esta segunda señal no aparecía respuesta inmune.18 (Figura 2)
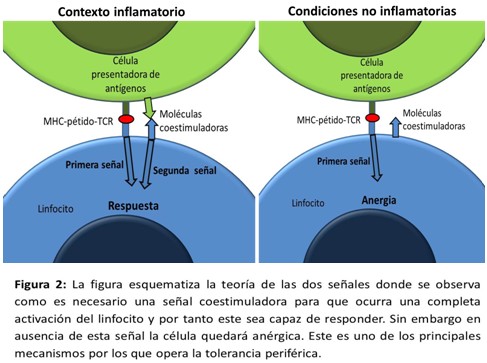
La segunda señal era generada por la célula presentadora de antígeno solo cuando esta se encontraba en un ambiente inflamatorio o ante antígenos foráneos, no expresando así esta segunda señal en el ambiente de lo propio. Esta teoría nos adelanta un paso para la teoría del peligro enunciada tres décadas después.
-La teoría de la red inmune
En 1974 Jerne propuso que los determinantes antigénicos configurados por las partes variables de las cadenas H y L de las inmunoglobulinas (idiotipos) constituían en sí un amplio repertorio de autoantígenos. Estos autoantígenos están en muy pequeña cantidad para ser destruidos por el sistema inmune, pero cuando un determinado clon es estimulado por su antígeno, el sistema inmune ya no podría ignorarlo y originaría unas respuestas contra ellos en forma de segundos anticuerpos (anti‑idiotipos) que encajarían con los primeros y tendrían además un efecto modulador que puede ser positivo o negativo. A este proceso se le denominó "teoría de la red de anticuerpos".19 (Figura 3)
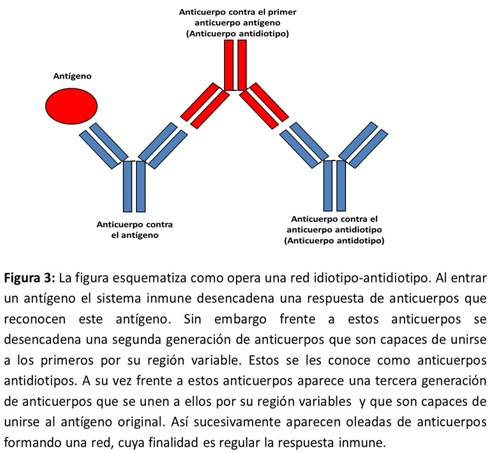
Esta teoría tiene como novedoso que por primera vez se reconoce la autorreactividad contra lo propio como un mecanismo fisiológico de regulación. Fue uno de los primeros hechos donde se reconoce la autoinmunidad como un fenómeno y sirvió de puente al cambio de paradigma de la inmunología desterrando el concepto de Erlich de horror autotoxicus.
II.-Década de los 90 y nuevas teorías del sistema inmune
Ya en la década de los 90 se acepta la autorreactividad natural como un fenómeno observado en la inmunología, tal es el caso de la presencia en individuos sanos de autoanticuerpos contra citocinas y la propia red idiotipo-antioidiotipo. De ahí que las nuevas teorías se enfoquen en cómo el sistema inmune logra la tolerancia basado en mecanismo activos, donde la respuesta tolerante es una modalidad de respuesta inmune que goza igualmente de especificidad y memoria; al igual que la respuesta inmune efectora. Por tanto para que aparezca una enfermedad autoinmune no basta con la presencia del clon autorreactivo sino que es necesario que exista un fallo en los mecanismos de regulación que evitan la activación de este clon.
-Teoría del homúnculo inmunológico
Esta teoría, plateada por Cohen en 1991, preconiza que en el sistema inmune existe una representación de todos los órganos y sistemas del cuerpo frente a los cuales se generan autoanticuerpos con fines regulatorios.20 Esta teoría por primera vez plantea el papel fisiológico que puede jugar la autorreactividad dentro y fuera del sistema inmune, siendo esto una visión distinta de la planteada por las teorías anteriores.21,22
-Teoría del peligro
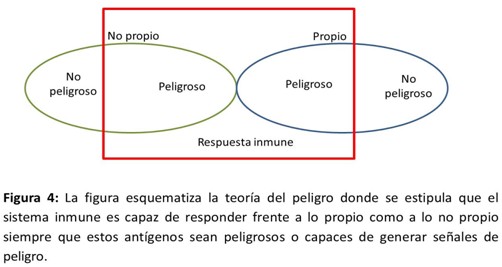
La teoría del peligro, postulada por Polly en 1994, plantea que los componentes propios podían desencadenar respuesta inmune si eran capaces de producir daño, mientras que componentes no propios pueden ser tolerados si estos no inducen daño.23
En esencia la teoría plantea que el problema no es ser propio o extraño sino la capacidad de producir daño tisular lo que determina si se produce o no una respuesta efectora. Para sustentar esta teoría los autores se basan en que el sistema inmune innato, a través de una serie de receptores de reconocimiento de patrones moleculares asociados a daño era el que daba la “instrucción” al sistema inmune adaptativo.24,25
Esta teoría explica, en parte, algunos de los mecanismos desencadenantes de las enfermedades autoinmunes, así cuando se presentan antígenos propios en un contexto de daño tisular aparece una respuesta inmune frente a estos componentes que desencadenan la respuesta autoinmune que concluye con más daño tisular y por tanto mayor intensidad de esta respuesta. Basados en esta teoría es que se puede justificar el fenómeno de autoperpetuación y empeoramiento progresivo de gran parte de las enfermedades autoinmunes. (Figura 4).
-Teoría de la tolerancia dominante
La teoría de la tolerancia dominante plantea que en individuos saludables existen células autorreactivas capaces de producir autoinmunidad y también existen células T reguladoras naturales que activamente mantienen la tolerancia. Esta teoría, como paso de avance, hace alusión al papel de la regulación en periferia de los clones autorreactivos26 y destaca en especial el papel de las células dendríticas (CDs) en el momento de iniciar la respuesta tolerante o efectora27-29 y el de las células T reguladoras al momento de iniciar y mantener esta respuesta inmune.30,31 Con la teoría de la tolerancia dominante se asume la tolerancia como un fenómeno activo, dotado de especificidad y memoria que no se diferencia de la clásica respuesta efectora en el cual intervienen los componentes del sistema inmune innato y adaptativo.
II.-Tendencias actuales
En la actualidad hay evidencia del papel de las células dendríticas en el establecimiento de la respuesta inmune; donde participan distintas subpoblaciones de CDs (CDs1, CDs2, CDs plasmocitoides, CDsCD4+, CDsCD8+) así como hay células reguladoras que también intervienen en el proceso de establecimiento o no de una respuesta inmune efectora o tolerante.32,33 De ahí que hoy en día se hable del papel de la regulación del sistema inmune en el establecimiento o no de la tolerancia como parte de una teoría compleja de redes,34 que se basa en un intrincado enrejado de interacciones celulares y moleculares. Las enfermedades autoinmunes aparecen ante un complejo de fenómenos que involucran fallos en estos mecanismos de control, unidos a pérdida de la tolerancia central y periférica; de ahí que el reducir la tolerancia a una teoría es reducir la magnitud del fenómeno, el cual descansa en una base multifactorial.
CONCLUSIONES
El sistema inmune opera bajo complejas interacciones celulares que le permiten “decidir” cuando reaccionar con una respuesta inmune efectora y cuando con una tolerante. Tratar de explicar cómo operan estos mecanismos ha sido el problema fundamental de la inmunología. Desde los inicios del pasado siglo numerosas teorías han sido enunciadas, cada una con aciertos y desaciertos; sin embargo han aportado un granito de arena a nuestros conocimientos actuales. Hoy en día se sabe que para que ocurra una respuesta efectora es necesario la existencia de señales de peligro y daño tisular, acompañado de un antígeno con propiedades inmunogénicas, que debe ser reconocido por un clon específico de linfocitos el cual se expande y se obtienen células hijas con igual especificidad, pero con afinidad mayor que las células vírgenes. También se sabe que en periferia existen clones autorreactivos los cuales tienen un papel fisiológico y son constantemente regulados por mecanismos de tolerancia periférica para evitar el desarrollo de enfermedades autoinmunes. El sistema inmune es complejo y diverso tanto como es la diversidad de antígeno al que se expone; es por eso que llegar a comprenderlo del todo es un reto al entendimiento humano.
